
2018年12月号 [Vol.29 No.9] 通巻第336号 201812_336001
熱帯林内の微環境変化とその生態的影響を追う —マレーシア熱帯林における実生と稚樹の長期観測—
1. はじめに
東南アジアの熱帯林は、世界の熱帯林の中でも高い生物多様性を持つばかりでなく、気候変動緩和にも重要な役割を果たしています。しかし、商業伐採や土地利用転換などによって、熱帯林は急速に減少、劣化しています。熱帯林の保全と持続可能な利用のためには、森林更新と植物環境応答のメカニズムを把握し、気候変動の緩和および適応の可能性をさらに深く理解することが急務です。
熱帯林の林床には、次世代の熱帯林を担う多様な実生[1]や稚樹[2]が生育しています。しかし、林冠部が高い位置にあり、複雑な階層構造を持つ熱帯林の林床は非常に暗く、実生や稚樹にとって重要な光資源(光合成有効光量子束密度:以下PPFD)が大きく制限されています(Yamada et al. 2014)。一方で、限られた光資源とは対照的に、CO2は下層植物にとっては十分にあります。閉鎖された熱帯林内では、土壌から排出されるCO2(土壌呼吸)によって、林床付近でCO2濃度が高くなる傾向があるためです(Ohkubo et al. 2008)。このような熱帯林内の光環境やCO2、温度などの微環境は、長期的な気候変動によって変化し、熱帯林の更新に影響することが予想されます。しかし、その影響に関する情報は非常に限られているのが現状です。
そこで我々は、熱帯植物におよぼす長期的な気候変動の影響を把握するために、東南アジア低地熱帯林において、微環境、植物の成長、フェノロジー(展葉、開花、落葉などの生物活動の季節性)、種や個体数の変化について長期観測を展開してきました。ここでは、マレーシア半島部のパソ森林保護区を拠点として、2011年から実施してきた、熱帯低地生態系の長期観測について紹介させていただきます。
2. 観測方法の詳細と観測結果
(1) 調査地の設置
我々は、パソ森林保護区に9カ所の観測プロットを設置し(図1)、林床の微環境を観測するとともに、実生と稚樹の個体数、フェノロジー、個体成長、種多様性に関する追跡調査を行っています。また、CO2濃度の上昇が実生や稚樹の生理機能特性に及ぼす影響についても、携帯型光合成測定装置(米国Li-Cor社のLI-6400)による現地計測を実施しています。
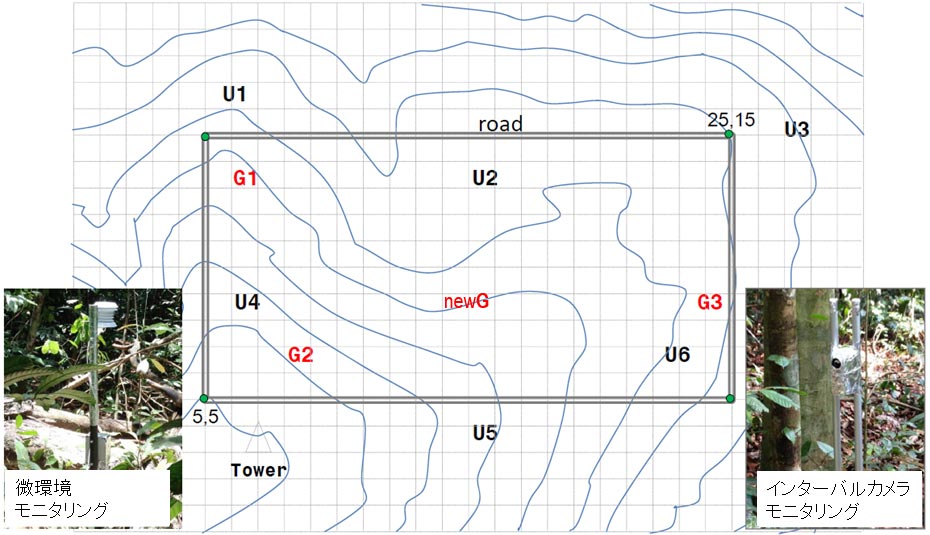
図1パソ森林保護区に設定した9カ所のプロット位置図。プロットは、2015年に倒木などによって林冠の開けた林床(ギャップ)に3カ所(G1〜G3)、林冠の閉じた暗い林床に6カ所(U1〜U6)に設置しました。また、new Gは2017年に確認された新しいギャップを示します。各プロットでは、気温、大気湿度、土壌水分、地温などの微環境モニタリングと自動撮影カメラによる実生フェノロジーのモニタリングを行っています
(2) 林床の微環境
パソ森林保護区の林床では、日中7割以上が、光合成に適さない、暗い光環境(PPFDが16μmol photons m−2 s−1未満)でした。一方で、晴天時にはPPFDが2000μmol photons m−2 s−1を超える直射日光の一部が林冠内部へ浸透し、林床にサンフレック[3]となって差し込んでいました。また、9つ全ての観測プロットの中央で全天写真を毎年2回撮影(Nikon coolpix950、写真1)した結果、林冠の開空度はサイト間で17.3%から4.5%(2016年6月)の範囲で変化し、ギャップでは年次ごとに減少する傾向が明らかになりました。このような微環境の空間的不均一性と時間的変動が、林床植物の変化にどのように影響を及ぼすかについて検討しています。

写真1各プロットの中央で、視野角180°の魚眼レンズを用いて天頂に向かって全天を撮影した一例(2016年1月25日撮影)。写真の白色部分が、林冠の開けた部分を示します。写真上方が北になります。林床プロット(U2、左)とギャッププロット(G2、右)の林冠の平均開空度(Mean openness: %)と、光強度の相対散乱光(SOC: %)を算出しました。黄色線は、それぞれ夏至と冬至の太陽軌道を示します
(3) 実生と稚樹のフェノロジー
フェノロジーカメラで得られた植物群落の画像解析によって、時系列にそって実生や稚樹の展葉、成長、個体の生存動態などを明らかにすることができます(写真2)。例えば、赤丸で示した稚樹は、8月1日から2枚の新葉を約1ヵ月かけて展開したことが分かりました。また、横たわる倒木が朽ちていく様子が時系列的に確認できました。このようなデータを用いることで、新しい視点から熱帯林の動態を理解することが期待されます。

写真2インターバル機能付きのカメラ撮影によって、林床の植生と環境の変化を1時間間隔でモニタリングしています。ここでは、2015年8月1日から9月10日までの間に、約2週間ごとに撮影されたギャッププロット(G1)の画像を示しています。赤で囲まれた範囲に、新葉が展開する様子が認められます
(4) 実生と稚樹の追跡調査
植物種多様性調査のために、9つ全ての観測プロットに、1m × 1mのサブプロットを5つ設置しています。植生調査は、2015年から1年ごとに実施しており、各プロットにおけるサブプロット内に出現した、全ての実生と稚樹の位置とサイズ、個体の消長を毎年記録しています(写真3)。
2014年4月にマレーシア半島部で大規模な一斉開花が発生しました。この一斉開花によって供給された種子が発芽し、2015年に多くの実生が発生しました。9プロットをすべて積算した、2015年から2017年3年間の現存個体数は、899、705、598個、植物種数は150、139、138種でした。現存個体数と植物種数のいずれも、2015年以降、年次を追って減少する傾向が認められました(図2)。また、3年間の追跡調査によって、生残が種特異的であることが明らかになりました。例えば個体数が多かった種は、3年間で個体数を大きく減らす傾向を示しました。この個体数の変化が、どのような環境要因によるのかについては、更なるデータの蓄積が必要です。
今後も林床環境や稚樹の種構成のモニタリングを継続し、各環境要因と森林更新の関係性を解明していくことで、熱帯森林生態系の構造と機能に対し、気候変動がおよぼす影響を評価することが可能となります。

写真3パソ森林保護区林床での植生追跡調査の様子。1m × 1mの5つのサブプロットを各プロットに設置し、その中に出現した全ての個体にラベルを付けています。このラベルをもとに、個体の生存、成長などに関する追跡調査を、毎年8月上旬に実施しています
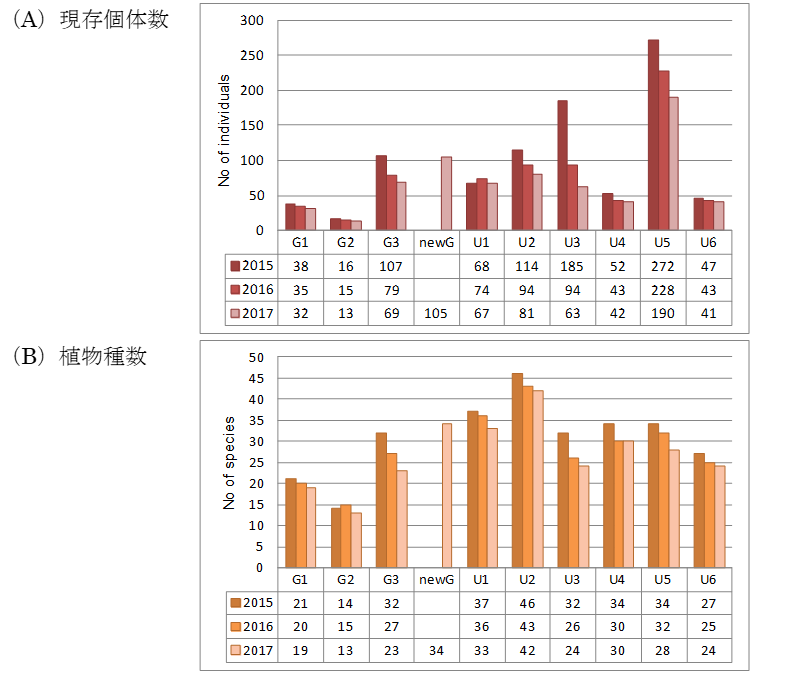
図2各プロットにおける現存個体数 (A)、植物種数 (B) の年次変化。林床プロット(U1〜U6)とギャッププロット(G1〜G3)における、2015年から2017年の3年間の年次変化を示しています。また、new Gは2017年に確認された新しいギャップを示します
(5) 光合成に対するCO2濃度の影響
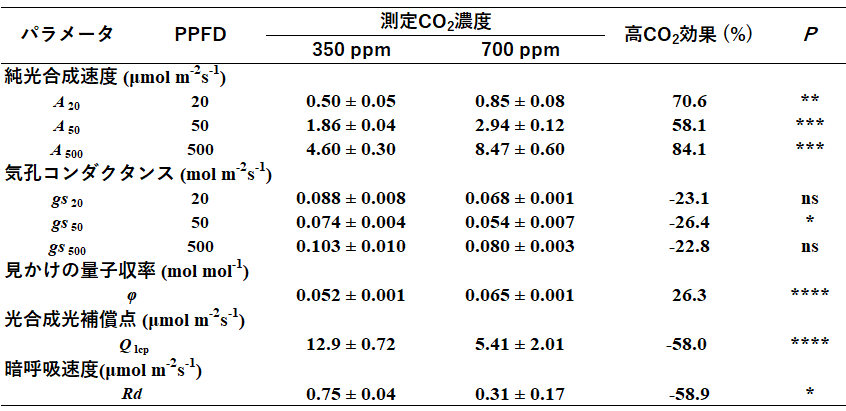
本研究では、パソ森林保護区で、環境変化に対する植物の生理生態学的応答メカニズムに関する研究も進めています。林床における高CO2環境は、そこに生育する植物の光合成光補償点(呼吸によるCO2放出と光合成によるCO2吸収のバランスがゼロになる光強度)を低下させるとされています(Hättenschwiler 2001)。光補償点が低いということは、すなわち弱光環境でもCO2を吸収できるということです(Liang et al. 2001)。また、暗い熱帯林林床に生育する植物においては、林冠の隙間から差し込む直射光、いわゆるサンフレックによりもたらされる短時間かつ強い光が重要だとされていますが、高CO2環境は、サンフレックに対する素早い応答を可能にし、光利用効率を上昇させているともされています(Tomimatsu and Tang 2012, 2016)。この2つの仮説を検証するために、東南アジア熱帯林における代表的な樹種である、フタバガキ科Dipterocarpus sublamelatus Foxw.の稚樹を用いた野外実験を行いました。光合成は、PPFDを人工的に制御しながらサンフレックを模した測定をしました(PPFDを20µmol photons m−2 s−1から500µmol photons m−2 s−1(30分間の照射)に急上昇させ、その後50µmol photons m−2 s−1(10分間の照射)に急低下させる)。また、PPFDに対する光合成速度の関係(光-光合成曲線)の計測から、光利用効率に関するパラメータ値を算出しました。光合成の測定は、熱帯林林床CO2濃度の変動幅の下限と上限を模した2つのCO2濃度(350ppm、700ppm)でそれぞれ実施しました。
その結果、D. sublamelatusの稚樹は、短期的なCO2濃度上昇によって、30分間の光照射で炭素吸収量を約2倍に増加させていました(図3)。この増加のうち、光照射に対する光合成反応の促進による効果が約7%であることが分かりました。また、短期的なCO2濃度上昇は、光合成光補償点を58%減少し、見かけの光量子収率を26%上昇させていました(表1)。これらの結果は、上記2つの仮説を支持しています。したがって、熱帯林内の短期的なCO2濃度上昇は、暗く安定した光環境とサンフレックなどの強光環境の両方で光利用効率を上昇させ、林床部の暗環境でも、植物によるCO2吸収量を補償していることが示唆されました(Tomimatsu et al. 2014)。
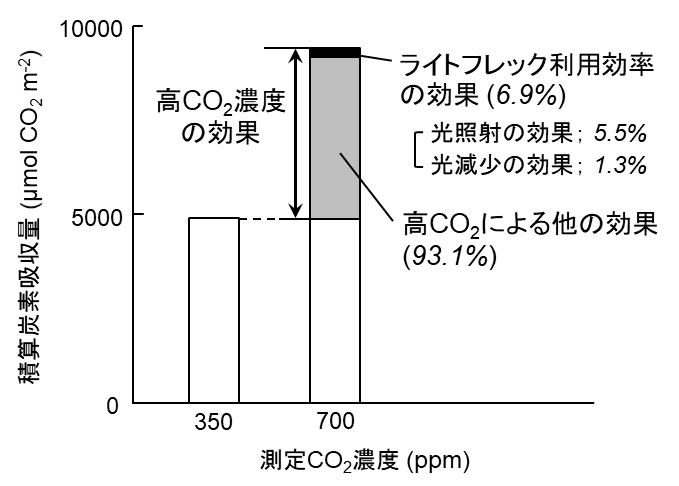
図3強光を30分間照射した時の積算炭素吸収量。サンフレックを模した強光(ライトフレック)を30分間照射したときに、光合成によって吸収されるCO2の積算量を示しています。図中の「高CO2による他の効果」には、「光合成基質としてのCO2施肥効果」と「定常光条件下での光利用効率の向上(見かけの量子収率や光補償点など)」があります。ライトフレック利用効率は、光強度変化に対する光合成反応速度の変化による効果を示しています
表1光定常状態での光合成の各種パラメータ値。純光合成速度と気孔コンダクタンスは、3つの光定常条件(PPFD:20、50、500µmol photons m−2 s−1)において、2つのCO2濃度(350ppm、700ppm)で測定しました。また、見かけの量子収率、光補償点、暗呼吸速度は、PPFDに対する光合成速度の関係(光-光合成曲線)から推定しました。また、CO2施肥にともなう変化率を、高CO2効果として算出しました
(Pは一元配置分散分析によるCO2濃度間の有意差を示す検定値であり、*の数が大きいほど有意であることを示します。*、**、***、****はそれぞれP値が < 0.05、< 0.01、< 0.001、< 0.0001であることを示しています)
3. まとめ
我々は、次世代の熱帯林を育む林床に注目し、実生や稚樹のフェノロジーのモニタリング、林床の光環境、CO2濃度と稚樹の光合成に関するメカニズムの研究を実施しています。
熱帯林における稚樹の種間個体数の消長に関する長期観測データの不足は、熱帯林更新の予測を妨げています。本研究により蓄積された基礎データを解析することで、植物種に依存した個体数の減少が示されました。また、熱帯林の林床植物が、林床の暗い環境下での物質生産に、高CO2濃度を有効利用できることを解明しました。これらの研究結果は、森林生態系に関する温暖化影響評価の高度化に貢献することが期待されます。今後は、微環境変化が植物機能におよぼす影響の評価に関する研究を掘り下げるとともに、微環境と植物成長のモニタリング結果を、多角的に解析する予定です。
参考論文
- Hättenschwiler S. (2001) Tree seedling growth in natural deep shade: functional traits related to interspecific variation in response to elevated CO2. Oecologia 129:31-42.
- Liang N., Tang Y. & Okuda T. (2001) Is elevation of CO2 concentration beneficial to seedling photosynthesis in the understory of tropical rain forests? Tree Physiology 21:1047–1055.
- Ohkubo S., Kosugi Y., Takanashi S., Matsuo N., Tani M. & Abdul Rahim N. (2008) Vertical profiles and storage fluxes of CO2, heat, and water in a tropical rainforest at Pasoh, Peninsular Malaysia. Tellus 60B:569-582.
- Tomimatsu H. & Tang Y. (2012) Elevated CO2 differentially affects photosynthetic induction response in two Populus species with different stomatal behavior. Oecologia 169:869-878.
- Tomimatsu H., Iio A., Adachi M., Saw L.G., Fletcher C. & Tang Y. (2014) High CO2 concentration increases relative leaf carbon gain under dynamic light in Dipterocarpus sublamellatus seedlings in a tropical rain forest, Malaysia. Tree Physiology 34:944-954.
- Tomimatsu H. & Tang Y. (2016) Effect of high CO2 levels on dynamic photosynthesis: carbon gain, mechanisms, and environmental interactions. Journal of Plant Research 129:365-377.
- Yamada T., Yoshioka A., Mazlan H., Liang N. & Okuda T. (2014) Spatial and temporal variations in the light environment in a primary and selectively logged forest long after logging in Peninsular Malaysia. Trees 28:1355-1365.
研究助成
本研究は、国立環境研究所国際環境研究事業戦略調整費「アジア熱帯生態系における生態系機能および人為的攪乱や気候変動による影響の解明に向けた研究推進を目的とするパソ観測研究拠点の強化」、環境省地球一括研究「(環1351)アジア陸域の指標生態系における温暖化影響の長期モニタリング研究」および国立環境研究所28年度所内公募研究 (A)「東南アジア熱帯林における高解像度3次元モニタリングによる生物多様性・機能的多様性の評価手法の開発」の支援により行われました。
脚注
- 実生 (seedling): 発芽し定着して間もない幼植物のことで、おおよそ高さ30cm未満の個体とする調査例が多い。
- 稚樹 (sapling): 実生が成長して次のライフステージに達しているが、成木のステージには達していない植物のことで、高さ30cm以上でDBH(胸高直径)5cm未満の個体とする調査例が多い。
- サンフレック (sunfleck): 植物群落内のように林冠や高茎草本によって被陰された場所にときおり到達する太陽直射光のことで、陽斑ともいい、いわゆる木漏れ日のことである。

